
イネは世界3大作物で、日本における主食として重要な作物です。稲作関係の内容の授業などは、アグリビジネス学科では永澤が、FCでは保田が担当しています。しかし永澤の専門はイネの遺伝学で、保田の専門は水田雑草(特にヒエ)です。しかし1プロでは、どちらも水田稲作に関する卒論指導を行っています。そこで1プロのイネ関連の研究分野には3分野あるのが現状となっています。以下にそれぞれの分野の研究紹介しておりますのでご覧ください。また、イネや水田雑草に関して研究したい学生さんがおられましたら、ご相談ください。
水稲直播き栽培は、育苗および田植えが省略できるため、省力的で、かつ大規模生産に適応した栽培技術として注目されています。しかし、イネの発芽や苗立ちの不安定さ (写真)、雑草制御の難しさ (除草剤2~3回+手取り除草)、収量の低さ・不安定さ、鳥害 (カモ、スズメ)などによって、導入を敬遠する農家も多く、特に秋田県では普及があまり進んでいません。水稲直播き栽培の普及拡大のためには、それら問題を解決していく必要があります。
まずは、イネの発芽率や苗立ち率を高めるために、種籾の播種量や播種方法について研究を行っています。また直播栽培にも、水田に水を入れた状態で行う湛水直播であるか入れない状態で播種する乾田直播の区別、また湛水直播でも播く種子の加工方法や播種の仕方など様々な形態があります。そこでFCでは平成23年度から様々な直播栽培を行っており、1プロの卒業研究では、平成24年度に奥山雄太が「イネの湛水直播栽培および乾田直播栽培の生育・収量の比較」 (鉄コート種子を湛水で点播する湛水直播と乾直の栽培比較試験)を、平成25年度には「乾田直播栽培における異なる施肥条件があきたこまちの生育へ及ぼす影響」と題して高木裕也が乾直を、「あきたこまち鉄コーティング直播における種子コーティング材の検討及び栽培試験」と題して高松慧理奈が、鉄コーティング種子を用いた湛水直播栽培試験を行い、平成26年には「あきたこまち乾田直播栽培における播種方法の検討」を熊谷和樹がそれぞれ卒業研究として行った。平成27年には1プロの3年次の活動として乾田直播と移植栽培のイネの生長の比較を行っている。

イネの直播栽培での出芽状況。鉄コート湛水直播栽培での出芽。安定していない

乾田直播栽培でも出芽にばらつきが見られるが、雑草が湛水直播よりも多くかつ大きくなり易く、問題
イネの全塩基配列の決定は日本が主導した国際的なイネゲノムプロジェクトで行われ、3大穀物の中では最も早く2002年に解読終了が宣言されました。それまでもイネは日本での作物研究の主要な材料でしたが、全塩基配列の決定により、世界的にモデル植物としての地位を確たるものとしました。モデル生物は実用的でない場合がほとんどですが、イネはモデル植物でありしかも実用的な作物であることから、高等生物の研究対象として特異な地位を持っていると言えます。つまり、モデル植物としてイネを研究することで、イネを作っている遺伝子の働きを理解すれば、新しいイネをデザインすることが出来るようになり、実用的でもあるかもしれないと言うことです。そういった新しい作物育種の基盤的知見としてイネの基礎研究の成果が、何時か役に立つ事もあるでしょう。
残念ながら現状でイネをデザインするといったほどの大がかりで有用な遺伝的改変をすることは出来ません。恐らく私たちのイネに対する理解はそれほどに深くはないからなのでしょう。ゲノムはほぼ総て解読したが、それはゲノムに書きこまれてある文字を認識しただけで、その意味はまだ良く理解されていません。イネのゲノムの半分以上の遺伝子が生物学的に何を行って居るのか不明です。ゲノムの意味を調べることは遺伝子の働きを調べることですが、ある遺伝子がイネの中で何をしているのか知るには、その遺伝子を破壊した突然変異体を作り、得られた突然変異体がどんな異常を持っているか調べる事から始めます。もとの遺伝子は突然変異体で見られる異常を起こさないように働いていたはずです。そこから初めてどんな働きを持つ遺伝子があるのかが解り、イネを作っている遺伝的なプログラムの一部の意味するところが解るのです。実際には私たちは毎年1000系統程度の突然変異処理されたイネを圃場に展開し(写真1)、その中から「?」と思うような突然変異体を見つけ、その意外な働きを持った遺伝子を単離するという方法で、遺伝子とその機能を結びつけようとしています。
では最近とれた突然変異体を中心として穂の突然変異体を幾つか紹介しましょう(写真参照)。

(写真1)田植え直後の田圃の様子。系統を管理するための木札が沢山立ってるのと、小さいイネの苗が見える

(写真2)穂の突然変異体の写真。一番上から15-AS-185•15-AS-375•14-AS-437•14-AS-262•15-AS-286•12-TS-118、野生型の穂
写真2の一番上の穂は2次枝梗を全く作っていません。上から2・3・4番目の穂は2次枝梗の数が減少し、恐らくその結果、小穂(籾)の数が減っています。上から5番目の穂はやはり2次枝梗の数が減りますが、穂の根本に近い1次枝梗についた2次枝梗が特に無くなります。6番目は穂の根本の方の一次枝梗は残りますが、穂軸は途中からから無くなるので、一次枝梗の数は減ります。2次枝梗の数もかなり減少します。一番下が普通のイネの穂です。
これらの突然変異体の異常から、ほぼすべての2次枝梗の分化を正に制御する遺伝子、全般的に2次枝梗の数を制御している遺伝子、2次枝梗の数、特に一次枝梗の付け根側の制御をしている遺伝子、穂軸自身の分化を通して、一次枝梗を作っていく能力を制御する遺伝子など様々な遺伝子が穂の分枝が生じる過程に働いていることが解ります。これらの突然変異体のなかの幾つかは既知の遺伝子の突然変異(文献1、2)が原因ですが、幾つかの物はまだ遺伝子が同定されていません。これまでに報告のない遺伝子であることを期待しています。
穂の形態形成に関する突然変異体では、分枝パターン以外にも節の形成に関する突然変異等の解析を行っています。他にも花の形態に関する突然変異体(写真3)、節の形成に関する突然変異体(写真4)、種子の大きさに関する突然変異体(写真5)その他胚の大きさに関する突然変異(文献3)など、いろいろな変異体を対象としています。

(写真3)内穎も外穎のような形態を示す突然変異体
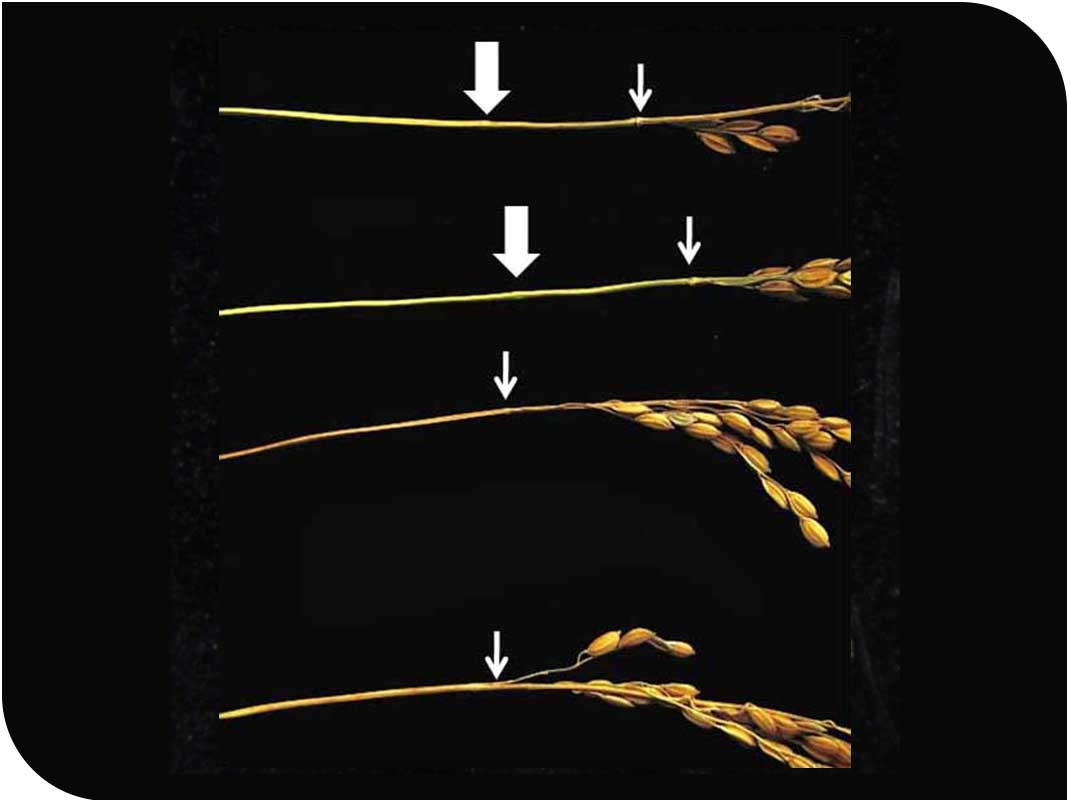
(写真4)節が本来無い場所に出来る(太い矢印)突然変異体。細い矢印は穂首節。上2本が突然変異体の穂で、下2本があきたこまちの穂

(写真5)種子が大きくなる突然変異体。A•E あきたこまち、B•F 11-AF-145、C•G 11-AF-192、D•H 11-AF-283
イネは年に1度しか育てられないこともあり、研究がなかなか進みません。是非やる気のある学生さんに手伝ってもらいたいところです。
【引用文献】
1. Komatsuet al. (2003). LAX and SPA: Major regulators of shoot branching in rice. Proc.Natl.Acad.Sci.USA. 100:11765-11770
2. abuchi et al. (2011). LAX PANICLE2 of Rice Encodes a Novel Nuclear Protein and Regulates the Formation of Axillary Meristems. Plant Cell 23:3276-3287
3. Nagasawa et al. (2013).GIANT EMBRYO encodes CYP78A13, required for proper size balance between embryo and endosperm in rice. Plant J. 75:592-605
イネの起源地は中国大陸です。日本には縄文時代晩期に伝播したと考えられています。イネの日本への伝播経路については諸説あるのですが、水利施設を伴う灌漑型の水田稲作(水稲農耕)については、縄文時代晩期の後半もしくは弥生時代初期に朝鮮半島南部を経由して九州北部に伝播したとする説が有力です。その水田に生育する水田雑草も、日本に自生していた野草から雑草化したものは少数であり、大多数は史前帰化植物(中国大陸から水田稲作とともに日本に侵入した)であると推定されてきました。しかし、史前帰化植物とされている水田雑草にも、稲作以前の更新世に存在したことが明らかになった種もあり、日本の水田雑草フロラの成立過程については再検討が必要になってきました。
日本での耕地面積の半分以上は水田であり、水田には耕地雑草の約40%にあたる191種の水田雑草が生育していることが確認されています。日本での水田雑草フロラは雑草フロラの中での重要な構成要素であり、その起点となる水田雑草フロラの成立過程の解明は、雑草生物学における重要課題であるとともに、初期の水田稲作の実態や雑草管理方法を推察していく上でも役に立ちます。
日本の水田雑草フロラの成立過程を解明するためには、初期の水田雑草の侵入や拡散の経路およびその時期を推定する必要があります。植物の侵入・拡散の経路や時期は、発掘された炭化種子や花粉を分析することの他に、種内の地理的変異を利用した系統地理学的研究によっても推定できます。例えば、水田雑草であるキツネノボタンでは、核型の地理的変異によって、農耕に伴って伝播(史前帰化植物)したのではなく、縄文時代以前に日本列島で生育していたと推定されました。
タイヌビエ(写真)は、イネ科ヒエ属の一年生水田雑草であり、日本、韓国、中国、インド北部、スリランカ、マレーシア、インドネシアなどのアジア地域やイタリア、フランス、北アメリカに分布します。タイヌビエは史前帰化植物であると考えられており、日本では北海道から沖縄県までの水田に広く分布しています。現在、日本各地からタイヌビエを収集し、その形態やDNAでの地理的変異を分析しています。少しずつではありますが、日本への侵入経路や拡散経路、稲作伝播との関連性がわかってきました。

水田に侵入するタイヌビエ

タイヌビエの穂

タイヌビエの小穂
【引用文献】
保田謙太郎・中山祐一郎:タイヌビエの小穂C型およびF型の日本国内での地理的分布、雑草研究 61.
保田謙太郎(2014)コラム:タイヌビエの変異からイネの伝播経路を探る『身近な雑草の生物学』(冨永達・根本編著)pp.86-87、朝倉書店
秋田県南秋田郡大潟村大潟6-5
TEL.0185-45-2858
FAX.0185-45-2415